Uncovering a mechanism of RNAP II oversampling in cell nuclei using combined SPT and FCS techniques.
In the environment of gene expression modulation, the disquisition and discovery of pivotal features come vital for recap controllers. It has been long accepted that RNA polymerase II( RNAP II) is signed to target genes by general recap factors during thepre-initiation complex( snap) assembly.
originally, RNAP II was observed to be statically deposited within clusters in the capitals of mammalian cells,co-localized with incipient RNAs 7, 8, andco-localized with invigorated RNAPII 7, 8. This led to the conception of stable recap manufactories.
Recent advancements in the field, still, reveal that RNAP II assembly is largely dynamic throughout its entire life cycle and can retain multiple RNAP II units, forming flash clusters that disassemble seconds after transcriptional exertion 9, 10, 11. How these clusters relate to the conception of recap manufactories or topologically associated disciplines( Boychicks) 12 and how they’re regulated over time remains unclear.
Our analysis has also unveiled the conception ofover-sampling by RNAPII1 through subunits. Grounded on these findings, we propose a model for flash recap regulation. Overall, our results exfoliate light on the significance of microscopy perspectives at different timescales to gain a comprehensive understanding of molecular dynamics in cells.
** dimension of Molecular Dynamics ** In recent decades, colorful microscopy perspectives have been developed to measure and/ or track molecular dynamics. Particularly, luminescence microscopy has been extensively employed to understand the dynamics of molecular unity, especially in the environment of spatiotemporal molecular processes, an essential element for understanding cellular function 16.
Measuring the dynamics of individual motes of both realities over time, at fast and slow spatial scales, is essential for understanding the dynamics of transcriptional events, including recap inauguration, along with molecular events in the nexus. For this purpose, we exercised the complementarity of luminescence correlation spectroscopy( FCS) high spatiotemporal resolution and single flyspeck shadowing( SPT) delicacy in original relegation measures, to claw deeper into RNAP II dynamics.
FCS relies on the dimension of oscillations in luminescence intensity due to the passage of fluorescent motes through the focal volume of a ray ray. Although the focal volume is small( about0.1 µm3), it corresponds to a subdomain that can accommodate numerous different molecular crowding countries or molecular relations, therefore forming an normal of several actions formerly 16.
also, such a change- grounded fashion is eyeless to static motes, a limitation that should be kept in mind 17. FCS is suitable for processes that do faster than 1 µm2/ s 19. also,super-resolution SPT ways have been developed in the once decade due to advancements in fluorophore and camera perceptivity, allowing the shadowing of individual motes in the range of times 20, 21.
Their flash nature frequently precludes representation as an normal of several actions, as seen by longer- track slow- moving realities, potentially furnishing a lesser representation of slow motes and less accurate estimates of fast motes, frequently eased by reducing SPT analysis 3.
In summary, the use of luminescence and camera perceptivity has enabled the development ofsuper-resolution SPT ways, which, together with FCS, allows for the shadowing of individual motes in a dynamic cellular terrain.
In this study, we first assessed the results attained through FCS( luminescence Correlation Spectroscopy) and SPT( Single flyspeck Tracking) ways with reference samples, in order to establish a standard for the use of these ways in understanding the geste of RPB1( RNA Polymerase II) in the nexus.
This helped us gain control over implicit impulses that could arise from different ways and different logical protocols used at different times. We argued that using experimental data rather of simulations was pivotal because it provides the most accurate description of real- life situations, considering the complications of live cells, walls, and benefactions.
We introduced a robust procedure to demonstrate adaptive conditions, which we applied to the analysis of RPB1 dynamics.
As a result, we showed that RPB1 dynamics in living cells can be described by at least four distinct dynamical modes within a range of0.1 to 15 µm ²/ s ³, 22, depending on the spatial objects conceivably related to RPB1- intermediated original target hunt, characterized by miscellaneous prolixity.
Results Before applying FCS and SPT ways for probing RPB1 dynamics, we first characterized their perceptivity in our setup, particularly grounded on the spread of fluorescent globules in different media with controlled density.
In order to replicate the observed clear spread values in the literature for protein prolixity in the nuclear terrain of cells, we named applicable criteria for FCS and SPT trials, videlicet between0.1 and 15 µm ²/ s ³, 22( see Accoutrements and styles for further details).
FCS dimension with ACFF Analysis for Calibrated Spread of globules FCS trials were conducted with fluorescent 40 nm periphery microspheres in water/ glycerol fusions with glycerol rates ranging from 0 to 80.
These combinations allowed us to demonstrate controlled prolixity portions in the range of 1 to 15 µm ²/ s( Table S1).
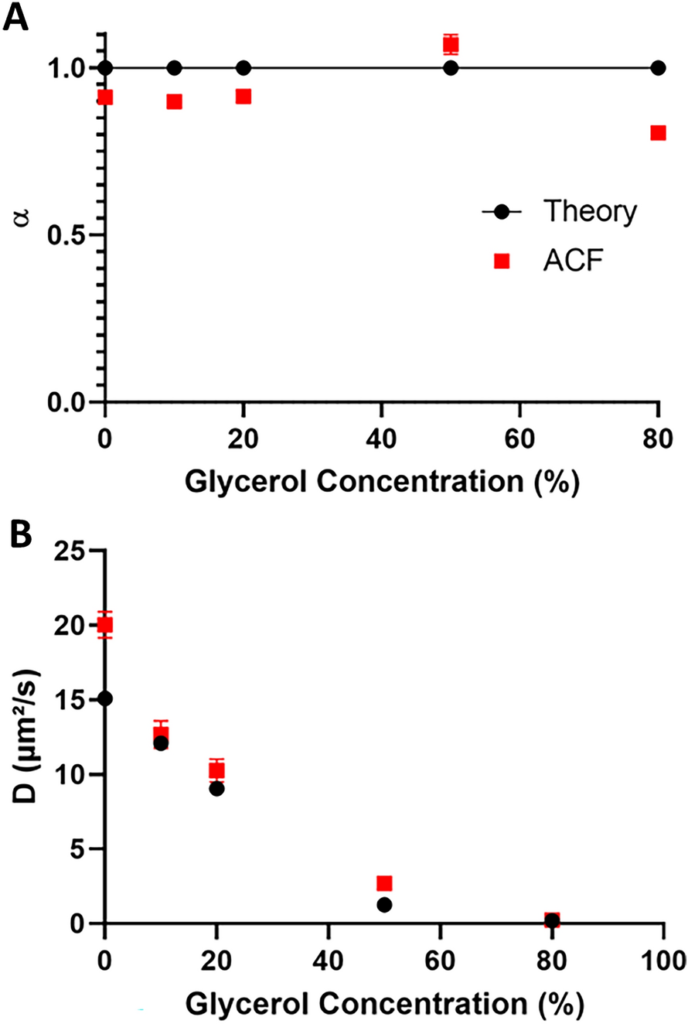
We chose to fit the autocorrelation angles produced by these trials to a miscellaneous model, generally used when studying molecular dynamics in living cells, similar as RPB1 dynamics in cellular capitals. We set up a good correlation between global literature values and our FCS experimental values( Figure 1 and Table S1). Up to 50 glycerol content in water, the spread of globules showed a remarkably close- to- concinnity anomalous measure( between0.96 and1.02) from ACFF analysis, as anticipated for Brownian prolixity( Figure 1A).
Only in the 80 glycerol- containing result did the blob spread show a significantly counterculturist value of0.70 ±0.02 with nascence( α), indicating miscellaneous spread, probably due to the high density of glycerol, which can disrupt complete unity as described in the literature preliminarily. also, we observed a slight overestimation in the spread measure values in our results compared to literature values, particularly for blob spread in pure water( Figure 1B).
This distinction in estimated prolixity portions might arise from minimum crimes, as reported preliminarily and outlined in accoutrements and styles. In summary, in our FCS setup, we can estimate prolixity portions in the range of 1 to 15 µm ²/s. Characterization of Bead Spread by SPT Analysis To compare measures by FCS and SPT, we applied a analogous strategy for assaying blob spread using SPT with the reference globules.
We extended the blob periphery up to 100 nm to identify slower events and reduced the circumstance of slow events( see Accoutrements and styles for details).
We demonstrated SPT accessions with SPIM( Single Aeroplane Illumination Microscopy) mode29, a fashion generally used for nuclear imaging as it allows for precise visualization due to the use of largely inclined and laminated optic wastes.
We linked and minimized the presence of slow events by adding the blob size to 100 nm( see Accoutrements and styles for further details). Using the MTT( Multiple- Target Tracking) algorithm software30, we anatomized image sequences to determine and reconstruct flyspeck circles.
Since small deportations stabilize the estimation function for these techniques31, we named only tracks with further than 10 findings, which are more dependable.
With this arrestment, we attained an average number of findings more or equal to 20 for all conditions( Figure S1). In line with FCS analysis, line analysis was performed with a miscellaneous model. We attained an estimated exponent( α) veritably near to 1, harmonious with the anticipation that globules in the glycerol admixture should parade Brownian stir( Figure 2A).
As seen in FCS, the high glycerol( 80) content in the admixture showed a counterculturist α value of0.8, suggesting miscellaneous spread probably associated with a certain diversity in the admixture.
A expertise is locating specific flyspecks. The water/glycerol mixture, which ranged in glycerol content from 0 to 80, was used to resuspension the fluorescent microspheres. SPT accessions were performed at a frame rate of 100 hertz.( A) An asymmetrical measure α was handed as a function of the glycerol rate.
Experimental values( red places) can be compared to theoretical values( black blotches).( B) The prolixity measure was calculated in two ways attained from the pitches of TEE- MSD( Time Ensemble Average Mean Square Displacement) angles, expressed in log form, for 60 ms time intervals, while D60ms was calculated for each line during the first 60 ms. Its circumstance is indicated by a red forecourt.
These experimental values can be compared to theoretical values( black blotches). Both styles generated a unforeseen change in D for high D values.( C) D60ms and α distributions from different experimental conditions are shown in a log- gauged smatter plot.
A specificsub-population can be linked within the criteria of certain parameters( green, grandiloquent, and unheroic blotches) while some imbrication with others( unheroic, red, and blue blotches). Data come from at least one hundred measures per condition.
also, we calculated the prolixity measure Dα from the spatial autocorrelation of MSD, as mentioned in the former paragraph. likewise, since it’s assumed that, over short time intervals, patches bear like Brownian patches, we calculated the D60ms values up to its original value by considering the pitch of the vectorial relegation on a log scale for each line over the first 60 ms, as preliminarily done for community32.
Both styles attained slightly lower prolixity measure values compared to the theoretical values( Figure 2B), as preliminarily described and bandied in reference 33. Interestingly, the divagation of the D60ms values related to the Dα reference was twofold lower, so we chose the D60ms system for our posterior SPT analysis of RPB1 prolixity.
We observed that for prolixity measure values above 4 µm2/ s, D60ms and Dα values were lower than the anticipated theoretical values, suggesting that we had reached a table on the volume. therefore, this estimation phase shows that SPT allows the study of prolixity insub-populations of different amounts of spreading patches, over several orders of magnitude.
To assess this conception that our approach can distinguishsub-populations in a mixedsub-population, as is the case in cells, we constructed fusions of spreading angles within different glycerol fusions. We chose to use MSD( appertained to as H- MSD) histograms analysis, which allows carrying a comprehensive overview of the spread features in complex samples grounded on MSD analysis of each community32.
Without resorting to unsupervised machine literacy, we estimated the applicable k number of populations( ranging from 1 to 10) by assaying the BIC( Bayesian Information Criterion) and AIC( Akaike Information Criterion) as k diminishments in the function of AIC k.
We also calculated GMM( Gaussian Mixture Model) with this k to fit our distribution. We tested this system’s capability to distinguish and recover both the correct values and the correct rate for different fusions of colorful glycerol substances by incorporating data from different MFI accessions. constantly, BIC and AIC analysis indicated that the two- population model stylish described our different fusions( Figure 3A, B 20 and 50 glycerol conditions admixture).
We showed that the H- MSD/ GMM system allowed differencing else spread features insub-populations and recover both the correct value and rate for both( Figure S2 and Table S3). Logically, this system was pushed towards its limit when we mixedsub-populations that were veritably close to the upper limit of discovery, i.e., when the populations centered around3.8 and4.0 µm2/ s were intermingled, which were considered as a single population( Table S3D).”
[…] United Airlines flight attendants are planning to carry a kick on Thursday morning. They’re taking action and calling for immediate changes in the plant after times of grueling […]